Translate this page into:
Filaggrin mutations and the skin
Correspondence Address:
Dipankar De
Department of Dermatology, Venereology, and Leprology, Postgraduate Institute of Medical Education and Research, Chandigarh
India
| How to cite this article: De D, Handa S. Filaggrin mutations and the skin. Indian J Dermatol Venereol Leprol 2012;78:545-551 |
Abstract
Filaggrin is very important in the terminal differentiation of the skin and the formation of cornified envelope in the stratum corneum. Several mutations in the filaggrin gene have been identified in the last decade, mostly from the European countries. Loss of function mutations in the filaggrin gene results in reduced production of filaggrin, depending on the type and site of mutation. Such mutations in the filaggrin gene have been shown to be the most significant genetic risk factor for development of atopic dermatitis and undoubtedly has a role in the pathogenesis of ichthyosis vulgaris. Though there is theoretical possibility of association with hand eczema and allergic contact dermatitis; in clinical studies, the strength of these associations was not significantly strong. In this review, we have discussed the structure and function of filaggrin, basic genetics, type of mutations in filaggrin gene, and association of such mutations with different dermatoses.Introduction
Filaggrin (filament aggregating protein) is an important element in the formation of statum corneum and thus in terminal differentiation of epidermis. Keratohyaline granules found in the granular layer of the epidermis are predominantly composed of profilaggrin, a 400- kDa precursor protein for filaggrin. The profilaggrin gene (FLG) is located in 1q21, which represents a dense cluster of genes encoding proteins involved in the terminal differentiation of epidermis. In addition to FLG, this epidermal differentiation complex (EDC) also accommodates genes for other proteins like loricrin, involucrin, trichohyalin, small protein rich proteins and others. Profilaggrin is cleaved into 10-12 copies of 37 kDa filaggrin peptides, which rapidly aggregate the keratin filaments causing collapse of the granular cells to form flattened anucleus squames of stratum corneum. The condensed cytoskeleton is cross-linked by transglutaminases forming cornified cell envelop. Barrier function of the cornified cell envelop prevents water evaporating out of the skin in the atmosphere and noxious environmental elements from entering into and affecting the human skin. It is worthwhile to mention that though filaggrin is an intracellular protein, the barrier function imparted by filaggrin is due to paracellular phenomenon; abnormality in keratin filament organization, impaired loading of lamellar body contents and associated non-uniform extracellular distribution of secreted organelle contents, as well as abnormalities in lamellar bilayer architecture. Very recently, it has been observed that FLG mutations predict dose-dependent alterations in epidermal permeability barrier function. [1]
In addition to terminal differentiation and barrier function of the epidermis, filaggrin has several other beneficial roles to play. Upon degradation of filaggrin, several amino acids (alanine, pyrrolidone carboxylic acid, urocanoic acid) are produced that act as natural moisturizing factors in the stratum corneum, [2] and also reduce the skin surface pH. [3] Histidine released from filaggrin degradation acts as putative ultraviolet photoprotector. [4] S-100 calcium binding domain in the molecular structure of profilaggrin may have an additional role in calcium signaling. [5]
The FLG like other members of ′S-100′ fused type gene family consists of 3 exons and 2 introns. [6] The initiation codon for FLG is located in exon 2 while the bulk of profilaggrin is encoded in exon 3. Number of the filaggrin repeats may vary due to duplication of either repeat 8 or repeat 10. [7] Such a large number of repeats is rarely seen in other genes and thus is unique for FLG. [8] Please refer to [Figure - 1] for the schematic representation of the domain structure of FLG. Out of around 40 mutations described in the FLG thus far, most are observed in people of European ancestry. The common mutations in FLG observed in the European and Asian population have been mentioned in [Table - 1].
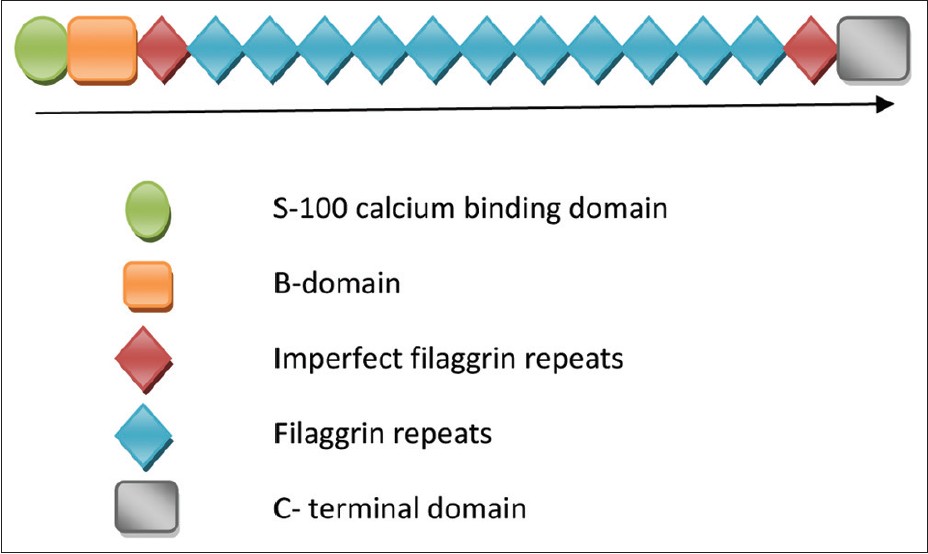 |
| Figure 1: Schematic diagram of domain structure of profilaggrin |

These include frame-shift or mis-sense mutations resulting in pre-mature termination of mRNA synthesis. The more proximal the mutation is towards N-terminal domain, the more severe is the deficiency in ultimate profilaggrin expression. Though more distal mutation allows limited expression of profilaggrin, practically no production of functional filaggrin occurs implying an important role of C-terminal domain in filaggrin processing. [9] These mutations are semi-dominant i.e. heterozygotes show incomplete penetrance and mild phenotype. [8] However, instead of analyzing individual FLG mutation phenotypes in isolation, if all known phenotypes of FLG mutation (ichthyosis, keratosis pilaris, AD, and palmar hyperlinearity) are considered together, penetrance is 100% in presence of 2 FLG mutations, 87.8% with one FLG mutation, and 46.5% in wild- type individuals. [10]
The expression of the filaggrin also depends on the type of inheritance of FLG mutation. The lowest expression occurs with homozygous FLG mutation (same mutation in both alleles) in any particular loci while the expression is normal in wild-type homozygotes (both alleles are normal). Intermediate level of expression occurs in patients who are heterozygous (mutation affecting only one allele) or compound heterozygous (both alleles of a gene harbor a mutation, but the mutations are different).
Recently, several new mutations has been described in Asian patients like in Korean, [11] Taiwanese, Japanese, and Singaporean Chinese [12] ichthyosis vulgaris patients. There was overlap between FLG mutations in different oriental population, probably due to their common ancestry till a certain time in the past. The overall differences in FLG mutations in different geographical location underscores the fact that one cannot use European FLG mutation data to screen Asian patients and thus information on FLG mutation should be obtained for each population. [12] Interestingly, in Pakistani ichthyosis vulgaris patients, both R501X [13] and 2282del4 [14] were found to be commonest in Europeans.
Filaggrin Mutation and Dermatoses
The significant role of FLG mutations in ichthyosis vulgaris and atopic dermatitis (AD) has been proven. Its limited role is found in contact dermatitis to nickel. Interestingly, FLG mutation has been found to exacerbate phenotypic expression of X-linked recessive ichthyosis. [15] In addition to mis-sense mutation of steroid sulphatase gene precipitating phenotypic features of XLRI, this reported patient had FLG mutation R501X. Owing to high population frequency of FLG mutations, FLG can act as possible genetic modifier in other genodermatoses. [16] Inheritance of FLG mutation in addition to mutations specific for the respective diseases is known to modify severity of pachyonychia congenita and alopecia areata. [17] Expression of filaggrin in other keratinizing epithelia like oral mucosa [18] and conjunctiva [19] may suggest that these organs can also be affected by FLG mutation. Keratoconus, however, was not found to be associated with FLG mutations. Of 89 keratoconus patients, only 5 were FLG mutation carrier whereas 43% patients had associated AD. [20] The theoretical possibility of association between FLG mutations and psoriasis arises because some genetic loci are shared by both AD and psoriasis in the EDC, and filaggrin expression is downregulated in psoriatic lesion while it is normally expressed in the uninvolved skin. However, Thyssen et al, in a meta- analysis concluded that the mutations R501X and 2282del4 are not causally associated with psoriasis. [21]
Filaggrin Gene Mutation and Ichthyosis Vulgaris
It is known that in ichthyosis vulgaris the granular layer is reduced or absent. Keratohyaline granules in addition to Odland bodies are major components of the granular layer, though the granularity is generally imparted by keratohyaline granules. Filaggrin being the major component of keratohyaline granules; the reduced expression is associated with reduced or absent granular layer. In addition, mRNA of profilaggrin in ichthyosis vulgaris patients has been found to be unstable with much shorter half- life. [22] R501X and 2282del4 mutations in FLG were found in pediatric patients with ichthyosis vulgaris who had abnormal skin barrier function. [9] Homozygous or compound heterozygous mutations R501X, and 2282del4 of the FLG has been shown as a cause of moderate to severe ichthyosis vulgaris. These mutations have combined allelic frequency of around 4%, explaining high incidence of ichthyosis vulgaris. This translates into the fact that 1 in 25 individuals will be heterozygous for FLG mutation while 1 in 2500 will be homozygous. In addition to ichthyotic skin, the phenotypic characteristics of ichthyosis vulgaris includes keratosis pilaris, hyperlinear palms, and very fine scales prominent on lower abdomen, arms, and legs. Heterozygotes have an intermediate phenotype with palmar hyperlinearity and in some cases, fine scaling and/ keratosis pilaris, whereas homozygous or compound heterozygous patients have more severe ichthyosis. [9]
Filaggrin Gene Mutation and Atopic Dermatitis
Until around half a decade back, AD was considered to be a primary immunologic disease where upon sustained exposure to external allergens, susceptible individuals mounted Th 2 response and eczematous lesions. [23] This inflammatory response provoked an impairment of skin barrier function the so-called inside-outside hypothesis. In the recent past, several studies have determined the significant association of FLG mutation with atopic eczema. Rapidly growing body of evidence on this has shifted the concept towards outside-inside hypothesis, wherein primary barrier abnormality precipitates downstream immunologic abnormalities. [24] According to this hypothesis, Th 2 response results from sustained and efficient epicutaneous allergen penetration through impaired skin barrier.
If it is agreed that FLG mutation is causative in pathogenesis of AD, what happens in patients with ichthyosis vulgaris where the mutation is the same but the disease expression is different? Are there additional triggers that precipitate eczematous lesions? It is believed that the permeability barrier in epidermis is normally imparted by the lipid-enriched extracellular matrix of the stratum corneum and not by cornified cell envelope. [25],[26] However, Scharschmidt et al, have shown that filaggrin deficiency itself is sufficient for impaired epidermal extracellular barrier function. [27] Unprocessed profilaggrin may impede extracellular secretion of lamellar bodies, the source of extracellular lipids. Alternatively, reduced production of acidic metabolites of filaggrin may result in net increase in skin surface pH activating neutral pH dependent kallikreins with unfavourable downstream effects on the barriers. [28],[29] The elevated pH can also deactivate enzymes responsible for synthesis of ceramides, the major lipids in maintaining epidermal permeability barriers. [30] Filaggrin deficiency alone decreases the threshold for inflammation in mouse model after application of irritants or haptens. [27] Scharschmidst et al, also demonstrated a phenotype similar to AD following exposure to reduced concentrations of hapten, which is normally unable to elicit Th 2 response in normal mice. Therefore, it is possible that recurrent epicutaneous exposure to haptens required for development of AD phenotype in filaggrin-deficient mice may not occur in ichthyosis vulgaris cases. In another study, Fallon et al, provided evidence that antigen transfer through a defective epidermal barrier is the principal mechanism responsible for an elevated IgE and initiation of cutaneous inflammation with phenotype similar to AD in FLG-deficient mice. [31]
Filaggrin mutations are the strongest and most widely replicated genetic risk to the pathogenesis of eczema. [32] Between 18-48% patients of AD in different series bear one or the other FLG mutation. [33] That is particularly true in Europeans with severe disease. [34] In 2 recent large population-based studies involving British [35] and German children, [36] the role of FLG mutation in AD was analyzed. In the German study, FLG mutations were associated with more than 3-fold increased risk of AD. In a systematic review and meta-analysis assessing risk of allergic disorders in FLG mutation patients, the risk of development of AD was 2-fold in family studies and 4.78-fold in case-control studies. [34] Population attributable risk of AD from mutations R501X and 2282del4 was observed to be 11%. [37] Those AD patients who have FLG mutations appear to have earlier disease onset and more persistent course. In 2 different cohorts [38],[39] of AD patients who had persistence of disease beyond 2 years, the carrier frequency were 29.7% and 42%, respectively. Since skin barrier function is impaired since birth in FLG mutation patients, it is expected that AD in such patients will be of an early onset and persistent. [39],[40] FLG mutation also predisposes to a type of palmar hyperlinearity with more than 60% of those patients who had mutation manifested criss - cross hyperlinearity of the thenar eminence. [10]
What is the scenario in the significant proportion of AD patients who do not have any FLG mutation? Mutation studies in several ethnic groups outside Europe did not reveal any significant role of FLG mutation in AD. In a case-control study involving 110 AD/ ichthyosis patients of Ethiopian origin, only 1 patient had a novel heterozygous mutation and none had 1 of the 4 most common mutations observed in Europeans. [41] In 200 Italian AD patients, novel mutations hitherto undescribed in European population were observed though FLG mutations were not found causative for AD. [42] IL-4 and IL-13, important cytokines in Th 2 type response, downregulate filaggrin expression in keratinocytes. [43] Recently, it has been observed that IL-22, another important cytokine in AD, also downregulates filaggrin mRNA expression as a secondary event. [44] In addition, the expression of genes for enzymes involved in processing of profilaggrin and the generation of moisturizing factors from degradation of filaggrin was also impaired. Therefore, even though the debate on what comes first continues, filaggrin downregulation- primary or secondary, is very important factor in barrier impairment and thus in the pathogenesis of AD.
FLG mutation has also been proposed for atopic march. [37] In several studies, the associations between FLG mutation and AD, allergic rhinitis and bronchial asthma have been found to be significant. However, the associations with allergic rhinitis and bronchial asthma appear to get stronger when there is co-existing AD. [34],[35],[37] In the ETAC (early treatment of the atopic child) population, the risk of asthma in FLG mutation-positive patients was significantly high. [45] Rogers et al, have shown that FLG mutations can increase the risk of asthma in co-existence with eczema but not in isolation. [46] The interesting aspect here is that when bronchial mucosa does not express filaggrin, how FLG mutation imposes susceptibility to bronchial asthma? Cutaneous priming to aero-allergens in a susceptible individual with barrier abnormality may initiate long- lasting Th 2 type of response, followed by IgE formation and mast cell activation in different mucosal surfaces after an appropriate exposure. [47]
The role of FLG mutations R501X and 2282del4 in the susceptibility to eczema herpeticum have been assessed. Impaired barrier function due to FLG mutation predispose to increased viral penetration and replication in the skin. As Th 2 immune response down-regulates cutaneous innate immunity as well, the dual role of FLG mutation in barrier function impairment and upregulation of Th 2 immune response put such patients at increased risk of eczema herpeticum. [48]
Filaggrin Gene Mutation and Hand Eczema
Several factors influence the incidence of hand eczema. These include contact allergy, wet work, female sex, atopic dermatitis etc. Atopic dermatitis is probably the strongest risk factor for development of hand eczema. [49] As FLG mutation has been found in a significant proportion of AD patients in several European studies, it can be expected that FLG mutation may increase the risk of hand eczema. Moreover, impaired barrier function in FLG mutation patients, irrespective of atopic status, may increase the penetration of environmental noxious agents, and thus increase the risk of contact eczema including that of the hands. de Jongh et al, found patients with FLG mutation at an increased risk of chronic irritant contact dermatitis of the hands or of the forearms. [50] On the contrary, Lerbaek et al, in a twin study could not find an association. [51]
Molin et al, in their study involving 122 German patients with defined chronic hand eczema subtypes, observed no overall significant association between chronic hand eczema in general and FLG mutations R501X and 2282del4. However, heterozygous mutation in FLG may predispose to chronic hand eczema that is characterized by both allergic and irritant contact dermatitis. [52] In this cohort, chronic hand eczema was associated with excessive exposure to water and irritants. A heterozygous non-functional FLG mutation might be responsible for lower resistance to environmental damage. However, production from the normal allele will tend to be protective. When this protection is overwhelmed by exposure to water or irritants, epidermal barrier function gradually collapses allowing the penetration of allergens, and this explains combined presence of irritant and allergic contact dermatitis in heterozygous FLG mutation patients.
Thyssen et al, observed that presence of atopic dermatitis in those with FLG mutation increased the risk of self-reported hand eczema in the preceding 12 months while in absence of AD, FLG mutation did not predispose to risk of hand eczema. [49] Several attributes in the hand eczema patients like history of hand eczema ever, lower age at the debut of hand eczema, contact sensitivity to at least one allergen, contact sensitivity to fragrance mix, ethylenediamine, and neomycin increased through the 4 groups, i.e. no AD nor FLG mutation, no AD but positive FLG mutation, presence of AD but no FLG mutation, and presence of both AD and FLG mutation; the risk of hand eczema being highest in the group with both AD and FLG mutation. Another recent observation by Thyssen et al, is the increased risk of development of fissured dermatitis on hands and fingers in adults with FLG mutation. [53] They recommended the use of prophylactic emollient application in FLG mutation carriers to prevent the development of occupational hand eczema.
Filaggrin Gene Mutation and Contact Sensitivity
Filaggrin-deficient skin is susceptible for increased transepidermal water loss and easy penetration of environmental allergens. FLG mutations increase the risk of allergic type I sensitization to aeroallergens, allergic rhinitis, and asthma in combination with atopic dermatitis. [34] However, Carlsen et al, could not prove the susceptibility of FLG mutation patients to type 1 hypersensitivity to natural latex rubber proteins. [54]
No association was found between FLG mutation and nickel allergy, polysensitization, dermatitis, and hand eczema at the debut of contact dermatitis. [55] Nonetheless, patients with FLG mutation started having eczema at an earlier age, and there was a tendency towards longer disease duration.
Hypothetically, filaggrin-deficient persons should be more susceptible to development of nickel sensitivity. [56] Normally, histidine-rich proteins in the upper parts of the epidermis acts as chelating agent for nickel. Consequently, penetration of nickel through the epidermis should be enhanced in filaggrin-deficient patients, filaggrin being an important source of histidine in upper epidermis. However, studies did not find significant association between filaggrin-deficient status and increased chances of nickel sensitivity. Carlsen et al, found positive association between combined mutation of R501X and 2282del4 genes, and nickel sensitivity though the association was not found to be statistically significant. [55] Another study found a positive association only when a positive patch test to nickel sulphate appeared in the context of previous history of allergy to artificial jewelry. [57] In yet another study, an association was found when an adjusted analysis on women who were not ear-pierced was performed. [58]
Contact allergens are generally smaller than 500 Da and may penetrate the skin irrespective of skin barrier function. [59] Protein allergens are much larger. The smaller size of most of the contact allergens explains the lack of association between FNM and contact allergy in general and polysensitization in particular. [55]
Conclusions
FLG mutations have gained importance recently, and around 40 mutations in the FLG gene have been described thus far. The role of FLG mutation in ichthyosis vulgaris and its associated phenotypes have been proven beyond doubt. While it is considered to be the most important genetic risk factor for development of AD, the role is not probably significant outside of European population. Its role in development of hand eczema and contact dermatitis, more particularly nickel dermatitis, is mired into several caveats. Study on FLG mutation is yet to break ground in India.
| 1. |
Gruber R, Elias PM, Crumrine D, Lin TK, Brandner GM, Hachem JP, et al. Filaggrin genotype in ichthyosis vulgaris predicts abnormalities in epidermal structure and function. Am J Pathol 2011;178:2252-63.
[Google Scholar]
|
| 2. |
Rawlings AV, Harding CR. Moisturizers and skin barrier function. Dermatol Ther 2004;17 (Suppl 1):S43-8.
[Google Scholar]
|
| 3. |
Elias PM, Chol EH. Interactions among stratum corneum defensive functions. Exp Dermatol 2005;14:719-26.
[Google Scholar]
|
| 4. |
McLoone P, Simics E, Barton A, Norval M, Gibbs NK. An action spectrum for the production of cis-urocanoic acid in human skin in vivo. J Invest Dermatol 2005;124:1071-4.
[Google Scholar]
|
| 5. |
Markova NG, Marekov LN, Chipev CC, Gan SQ, Idler WW, Steinert PM. Profilaggrin is a major epidermal calcium-binding protein. Mol Cell Biol 1993;13:613-25.
[Google Scholar]
|
| 6. |
Gan SQ, McBride OW, Idler WW, Markova N. Steinert PM. Organization, structure, and polymorphisms of the human profilaggrin gene. Biochemistry 1990;29:9432-40.
[Google Scholar]
|
| 7. |
Sandilands A, Smith FJD, Irvine AD, McLean WHI. Filaggrin's fuller figure: A glimpse into the genetic architecture of atopic dermatitis. J Invest Dermatol 2007;127:1282-4.
[Google Scholar]
|
| 8. |
Smith FJD, Irvine AD, Terron-Kwiatkowaski A, Sandilands A, Campbell LE, Zhao Y, et al. Loss-of-function mutation in the gene encoding filaggrin cause ichthyosis vulgaris. Nat Genet 2006;38:337-42.
[Google Scholar]
|
| 9. |
Sandilands A, Terron-Kwiatkowaski A, Hull PR, O'Regan GM, Clayton TH, Watson RM, et al. Comprehensive analysis of the gene encoding filaggrin uncovers prevalent and rare mutations in ichthyosis vulgaris and atopic eczema. Nat Genet 2007;39:650-4.
et al. Comprehensive analysis of the gene encoding filaggrin uncovers prevalent and rare mutations in ichthyosis vulgaris and atopic eczema. Nat Genet 2007;39:650-4. '>[Google Scholar]
|
| 10. |
Brown SJ, Relton CL, Liao H, Zhao Y, Sandilands A, McLean WHI, et al. Filaggrin haploinsufficiency is highly penetrant and is associated with increased severity of eczema: Further delineation of the skin phenotype in a prospective epidemiological study of 792 school children. Br J Dermatol 2009;161:884-9.
[Google Scholar]
|
| 11. |
Kim EJ, Jeong MS, Li K, Park MK, Lee MK, Yoon Y. Genetic polymorphism of FLG in Korean ichthyosis vulgaris patients. Ann Dermatol 2011;23:170-6.
[Google Scholar]
|
| 12. |
Hsu CK, Akiyama M, Nemoto-Hasebe I, Nomura T, Sandilands A, Chao SC, et al. Analysis of Taiwanese ichthyosis vulgaris families further demonstrates differences in FLG mutations between European and Asian populations. Br J Dermatol 2009;161:448-51.
[Google Scholar]
|
| 13. |
Samdani AJ, Naz N, Ahmed N. Molecular studies of ichthyosis vulgaris in Pakistani families. J Coll Physicians Surg Pak 2010;20:644-8.
[Google Scholar]
|
| 14. |
Naz N, Samdani AJ. Detection of filaggrin gene mutation (2282Del4) in Pakistani ichthyosis vulgaris families. J Coll Physicians Surg Pak 2011;21:382-3.
[Google Scholar]
|
| 15. |
Liao H, Waters AJ, Goudie DR, Aitken DA, Graham G, Smith FJ, et al. Filaggrin mutations are genetic modifying factors exacerbating X- linked ichthyosis. J Invest Dermatol 2007;127:2795-8.
[Google Scholar]
|
| 16. |
Sandilands A, O'Regan GM, Liao H, Zhao Y, Terron- Kwiatkowski A, Watson RM, et al. Prevalent and rare mutations in the gene encoding filaggrin cause ichthyosis vulgaris and predispose individuals to atopic dermatitis. J Invest Dermatol 2006;126:1770-5.
et al. Prevalent and rare mutations in the gene encoding filaggrin cause ichthyosis vulgaris and predispose individuals to atopic dermatitis. J Invest Dermatol 2006;126:1770-5.'>[Google Scholar]
|
| 17. |
Gruber R, Wilson NJ, Smith FJ, Grabher D, Steinwender L, Fritsch P, et al. Increased pachyonychia severity in patients with concurrent keratin and filaggrin mutations. Br J Dermatol 2009;161:1391-5.
[Google Scholar]
|
| 18. |
De Benedetto A, Qualia CM, Baroody FM, Beck LA. Filaggrin expression in oral, nasal, and esophageal mucosa. J Invest Dermatol 2008;128:1594-7.
[Google Scholar]
|
| 19. |
Krenzer KL, Freddo TF. Cytokeratin expression in normal human bulbar conjunctiva obtained by impression cytology. Invest Ophthalmol Vis Sci 1997;38:142-52.
[Google Scholar]
|
| 20. |
Droitcourt C, Touboul D, Ged C, Ezzedine K, Cario-Andre M, de Verneuil H, et al. A prospective study of filaggrin null mutations in keratoconus patients with or without atopic disorders. Dermatology 2011;222:336-41.
[Google Scholar]
|
| 21. |
Thyssen J, Johansen J, Carlsen B, Linneberg A, Meldgaard M, Szecsi P. The filaggrin null genotypes R501X and 2282del4 seem not to be associated with psoriasis: Results from general population study and meta-analysis. J Eur Acad Dermatol Venereol 2012;26:782-4.
[Google Scholar]
|
| 22. |
Nirunsuksiri W, Zhang S-H, Fleckman P. Reduced stability and bi-allelic, co-equal expression of profilaggrin mRNA in keratinocytes cultured from subjects with ichthyosis vulgaris. J Invest Dermatol 1998;110: 854-61.
[Google Scholar]
|
| 23. |
Boguniewicz M, Leung DY. Atopic dermatitis. J Allergy Clin Immunol 2006;117:S475-80.
[Google Scholar]
|
| 24. |
Jung T, Stingl G. Atopic dermatitis: Therapeutic concepts evolving from new pathophysiologic insights. J Allery Clin Immunol 2008;122:1074-81.
[Google Scholar]
|
| 25. |
Elias PM, Menon GK. Structural and lipid biochemical correlates of the epidermal permeability barrier. Adv Lipid Res 1991;24:1-26.
[Google Scholar]
|
| 26. |
Elias PM. Stratum corneum defensive functions: an integrated view. J Invest Dermatol 2005;125:183-200.
[Google Scholar]
|
| 27. |
Scharschmidt TC, Man MQ, Hatano Y, Crumrine D, Gunathilake R, Sundberg JP, et al. Filaggrin deficiency confers a paracellular barrier abnormality that reduces inflammatory thresholds to irritants and haptens. J Allergy Clin Immunol 2009;124:496-506.
[Google Scholar]
|
| 28. |
Elias PM, Steinhoff M. "Outside- to- inside" (and now back to 'outside') pathogenic mechanisms in atopic dermatitis. J Invest Dermatol 2008;128:1067-70.
[Google Scholar]
|
| 29. |
Elias PM, Hatano Y, Williams ML. Basis for the barrier abnormality in atopic dermatitis: Outside-inside-outside pathogenic mechanisms. J Allergy Clin Immunol 2008;121:1337-43.
[Google Scholar]
|
| 30. |
Hachen JP, Man MQ, Crumrine D, Uchida Y, Brown BE, Rogiers V, et al. Sustained serine proteases activity by prolonged increase in pH leads to degradation of lipid processing enzymes and profound alterations of barrier function and stratum corneum integrity. J Invest Dermatol 2005;125:510-20.
[Google Scholar]
|
| 31. |
Fallon PG, Sasaki T, Sandilands A, Campbell LE, Saunders SP, Mangan NE, et al. A homozygous frameshift mutation in the murine filaggrin gene facilitates enhanced percutaneous allergen priming. Nat Genet 2009;41:602-8.
[Google Scholar]
|
| 32. |
O'Regan GM, Sandilands A, McLean WH, Irvine AD. Filaggrin in atopic dermatitis. J Allergy Clin Immunol 2008;122:689-93.
[Google Scholar]
|
| 33. |
Irvine AD. Fleshing out filaggrin phenotypes. J Invest Dermatol 2007;127:504-7.
[Google Scholar]
|
| 34. |
van den Oord RA, Sheikh A. Filaggrin gene defects and risk of developing allergic sensitization and allergic disorders: Systematic review and meta-analysis. BMJ 2009;339:b2433.
[Google Scholar]
|
| 35. |
Henderson J, Northstone K, Lee SP, Liao H, Zhao Y, Pembrey M, et al. The burden of disease associated with filaggrin mutation: A population-based, longitudinal birth cohort study. J Allergy Clin Immunol 2008;121:872-7.
[Google Scholar]
|
| 36. |
Weidinger S, O'Sullivan M, Illig T, Baurecht H, Depner M, Rodriguez E, et al. Filaggrin mutations, atopic eczema, hay fever, and asthma in children. J Allergy Clin Immunol 2008;121:1203-9.
et al. Filaggrin mutations, atopic eczema, hay fever, and asthma in children. J Allergy Clin Immunol 2008;121:1203-9.'>[Google Scholar]
|
| 37. |
Marenholz I, Nickel R, Ruschendorf F, Schulz F, Esparza-Gordillo J, Kerscher T, et al. Filaggrin loss-of-function mutations predispose to phenotypes involved in the atopic march. J Allergy Clin Immunol 2006;118:866-71.
[Google Scholar]
|
| 38. |
Weidinger S, Rodriguez E, Stahl S, Wagenpfeil S, Klopp N, Illig T, et al. Filaggrin mutations strongly predispose to early-onset and extrinsic atopic dermatitis. J Invest Dermatol 2007;127:724-6.
[Google Scholar]
|
| 39. |
Barker JN, Palmer CA, Zhao Y, Liao H, Hull PR, Lee SP, et al. Null mutations in the filaggrin gene (FLG) determine major susceptibility to early-onset atopic dermatitis that persists into adulthood. J Invest Dermatol 2007;127:564-7.
[Google Scholar]
|
| 40. |
Baurecht H, Irvine AD, Novak N, Illig T, Buhler B, Ring J, et al. Toward a major risk factor for atopic eczema: Meta-analysis of filaggrin polymorphism data. J Allergy Clin Immunol 2007;120:1406-12.
[Google Scholar]
|
| 41. |
Winge MC, Bilcha KD, Lieden AD, Shibeshi D, Sandilands A, Wahlgreen CF, et al. Novel filaggrin mutation but no other loss-of-function variants found in Ethiopian atopic-dermatitis patients. Br J Dermatol 2011;165:1074-80.
[Google Scholar]
|
| 42. |
Cascella R, Cuzzola VF, Lepre T, Galli E, Mosches V, Chini L, et al. Full sequencing of the FLG gene in Italian patients with atopic eczema: Evidence of new mutations, but lack of an association. J Invest Dermatol 2011;131:982-4.
[Google Scholar]
|
| 43. |
Howell MD, Kim BE, Gao P, Grant AV, Boguniewicz M, DeBenedetto A, et al. Cytokine modulation of atopic dermatitis filaggrin skin expression. J Allergy Clin Immunol 2009;124:R7-12.
[Google Scholar]
|
| 44. |
Gutowska-Owsiak D, Schaupp AL, Salimi M, Taylor S, Ogg GS. Interleukin-22 downregulates filaggrin expression and affects expression of profilaggrin processing enzymes. Br J Dermatol 2011;165:492-8.
[Google Scholar]
|
| 45. |
Muller S, Marenholz I, Lee YA, Sengler C, Zitnik SE, Griffioen RW, et al. Association of filaggrin loss-of-function- mutations with atopic dermatitis and asthma in the early treatment of the atopic child (ETAC) population. Pediatric Allergy Immunol 2009;20:358-61.
[Google Scholar]
|
| 46. |
Rogers AJ, Celedon JC, Lasky- Su JA, Weiss ST, Raby BA. Filaggrin mutations confer susceptibility to atopic dermatitis but not to asthma. J Allergy Clin Immunol 2007;120:1332-7.
[Google Scholar]
|
| 47. |
Hudson TJ. Skin barrier function and allergic risk. Nat Genet 2006;38:399-400.
[Google Scholar]
|
| 48. |
Gao PS, Rafaels NM, Hand T, Murray T, Boguniewicz M, Hata T, et al. Filaggrin mutations that confer risk of atopic dermatitis confer greater risk for eczema herpeticum. J Allergy Clin Immunol 2009;124:507-13.
[Google Scholar]
|
| 49. |
Thyssen JP, Carlsen BC, Menne T, Linneberg A, Nielsen NH, Meldgaard M, et al. Filaggrin null mutations increase the risk and persistence of hand eczema in subjects with atopic dermatitis: Results from a general population study. Br J Dermatol 2010;163:115-20.
[Google Scholar]
|
| 50. |
de Jongh CM, Khrenova L, Verberk MM, Calkoen F, van Dijk FJ, Voss H, et al. Loss-of-function polymorphisms in the filaggrin gene are associated with an increased susceptibility to chronic irritant contact dermatitis: A case-control study. Br J Dermatol 2008;159:621-7.
[Google Scholar]
|
| 51. |
Lerbaek A, Bisgaard H, Agner T, Ohm Kyvik K, Palmer CN, Menne T. Filaggrin null alleles are not associated with hand eczema or contact allergy. Br J Dermatol 2007;157:1199-204.
[Google Scholar]
|
| 52. |
Molin S, Vollmer S, Weiss EH, Ruzicka T, Prinz JC. Filaggrin mutations may confer susceptibility to chronic hand eczema characterized by combined allergic and irritant contact dermatitis. Br J Dermatol 2009;161:801-7.
[Google Scholar]
|
| 53. |
Thyssen JP, Ross-Hansen K, Johansen JD, Zachariae C, Carlsen BC, Linneberg A, et al. Filaggrin loss-of-function mutation R501X and 2282del4 carrier status is associated with fissured skin on the hands: results from a cross-sectional population study. Br J Dermatol 2012;166:46-53.
[Google Scholar]
|
| 54. |
Carlsen BC, Meldgaard M, Hamann D, Hamann Q, Hamann C, Thyssen JL, et al. Latex allergy and filaggrin null mutation. J Dent 2011;39:128-32.
[Google Scholar]
|
| 55. |
Carlsen BC, Johansen DJ, Menne T, Meldgaard M, Szecsi PB, Stender S, et al. Filaggrin null mutations and association with contact allergy and allergic contact dermatitis: results from a tertiary dermatology clinic. Contact Dermatitis 2010;63:89-95.
[Google Scholar]
|
| 56. |
Thyssen J, Carlsen B, Menn´e T. Nickel sensitization, hand eczema, and loss-of-function mutations in the filaggrin gene. Dermatitis 2008;19:303-7.
[Google Scholar]
|
| 57. |
Novak N, Baurecht H, Schafer T, Rodriguez E, Wagenpfeil S, Klopp N, et al. Loss-of-function mutations in the filaggrin gene and allergic contact sensitization to nickel. J Invest Dermatol 2008;128:1430-5.
[Google Scholar]
|
| 58. |
Thyssen JP, Johansen JD, Linneberg A, Menne T, Nielsen NH, Meldgaard M, et al. The association between null-mutations in the Filaggrin gene and contact sensitization to nickel and other chemicals in the general population. Br J Dermatol 2010;162:1278-85.
[Google Scholar]
|
| 59. |
Bos JD, Meinardi MM. The 500 Dalton rule for the skin penetration of chemical compounds and drugs. Exp Dermatol 2000;9:165-9.
[Google Scholar]
|
Fulltext Views
12,594
PDF downloads
3,367





![[Figure - 1]](#fig_ijdvl_2012_78_5_545_100518_u2.jpg){kind=link}
![[Table - 1]](#tbl_ijdvl_2012_78_5_545_100518_b1.jpg){kind=link}