Translate this page into:
Understanding itch: An update on mediators and mechanisms of pruritus
Correspondence Address:
Iffat Hassan
Postgraduate Department of Dermatology, STD and Leprosy, Govt. Medical College, University of Kashmir, Srinagar, Jammu and Kashmir
India
| How to cite this article: Hassan I, Haji MI. Understanding itch: An update on mediators and mechanisms of pruritus. Indian J Dermatol Venereol Leprol 2014;80:106-114 |
Abstract
Pruritus is the most common symptom secondary to skin diseases. Advances in the fields of neurobiology, immunology and physiology have made it possible for us to understand and unravel the deeper pathophysiological basis of pruritus. This review aims to update our current understanding of the mechanisms and mediators of pruritus. Special attention is paid to endogenous itch mediators particularly newly identified ones like endovanilloids, opioids, neurotrophins, cannabinoids, proteases and cytokines. Various theories explaining the peripheral encoding of itch are reviewed. Multiple neural pathways including the central itch pathways as well as supraspinal processing of itch and brain areas involved in pruritus are highlighted. Apart from peripheral itch mediators, spinal neural receptors are also involved in control of itch and should form part of the development of a novel antipruritic strategy. Further studies are required to fill the lacunae in our current understanding of the pathophysiology of pruritus.Introduction
In 1660 Samuel Haffenrefer proposed that itch be defined as an "unpleasant sensation that elicits the desire or reflex to scratch". [1],[2] However this definition is not entirely satisfactory because of the subjective nature of the term "unpleasant". [3] Furthermore, in many instances subjects rub instead of scratching in response to itch, as is quite commonly the case with patients of urticaria. [4] Itch may be regarded, in a teleological sense, as a physiological defense mechanism to keep at bay harmful external agents. [5] Itch is a predominant symptom of many cutaneous disorders like atopic eczema and various systemic diseases like liver failure and end-stage renal disease (ESRD), and certain neurological and psychiatric diseases as well. Itch, especially when chronic, is one of the most distressing sensations and can substantially impair the quality of life. [6]
Advances in dermatological and neurobiological research have lead to a significant unravelling of the pathophysiological basis of pruritus. Despite being the standard treatment for itch, antihistamines are not effective in lessening pruritus in various cutaneous and systemic diseases; [7] therefore, a better understanding of itch assumes a crucial importance from a therapeutic point of view.
Neuronal Pathways of Pruritus
Many theories have been proposed to elucidate the encoding of itch; the most important ones are as follows.
Intensity Hypothesis
For long itch has been considered as a sub-modality of pain. Intensity hypothesis postulates neurons that are activated by both painful and pruritogenic stimuli, but weaker activation of nociceptive receptors results in itch. [8] However, recent evidence against this theory is accumulating and suggests that the neurophysiology of itch is much more complex than initially thought. For example, when algogens are applied topically in lower concentrations it results in low intensity pain and not pruritus. [5]
Also, electrocutaneous stimulation of human afferent nerves revealed that an increase in the frequency of stimulation leads to more intense pruritus or pain without any qualitative change in the sensation from one modality to the other. [9]
Labeled Line or Specificity Theory
Microneurography experiments conducted in 1997 by Schmelz et al.,[10] identified afferent C-nerve fibers with slender axons and lot of terminal branching. These fibers were histamine sensitive but insensitive to mechanical stimuli. The detection of these low velocity fibers distinct from nociceptive fibers provided support for the labeled line theory which hypothesizes discrete and mutually exclusive afferent fibers for detecting either itch or pain. [11]
To detect whether these histamine-sensitive C fibers are represented centrally, Andrew and Craig [12] conducted electrophysiological recording in cats which revealed a subset of spinothalamic tract (STT) neurons in lamina I responsive to histamine but unresponsive to thermal and noxious stimulation as well as to the algogen, mustard oil. This strongly suggested that specific neuronal pathways are involved at both peripheral and central levels in mediating itch, thus supporting the labeled line theory.
However, it has been observed that in primates, histamine-sensitive STT neurons respond strongly and invariably to capsaicin, arguing against the labeled line theory. [13]
Furthermore, a study by Schmelz et al.[14] found that C fibers that were initially believed to be itch-specific also responded to capsaicin.
The Selectivity or Occlusion Model
This model combines aspects of both the preceding theories. It postulates that pruritoceptors are part of a larger set of nociceptors, the activation of the whole group elicits pain while activation of the itch-selective subset elicits itch. [11],[13]
In summary, it is still not entirely clear if the neural pathways for itch and pain are completely independent or not.
Multiple Neural Pathways for Itch Induction
Oral antihistamines are ineffective in the treatment of many types of pruritus, [7] suggesting a role for non-histaminic pathways in the sensation of pruritus. In the primate STT, distinct non-histaminergic C-nerve fibers transmitting itch have been discovered; these fibers are activated by cowhage (pruritogenic spicules which cover the pod of Mucuna pruriens, a tropical legume.). [15]
In a study by Namer et al., [16] cowhage spicules resulted in pruritus only in mechano-responsive C fibers (CM) but not in mechano-insensitive C fibers (CMi). This was in contrast to histamine which induced itch only in mechano-insensitive fibers (CMi). The study concluded that cowhage and histamine induce itch by activating separate non-overlapping groups of C fibers. Cowhage-induced itch does not produce flare and is not abolished by topical antihistamine treatment, however desensitization with topical capsaicin abolishes it, indicating that signaling of cowhage-induced itch occurs via a group of afferent nerve fibers sensitive to capsaicin. [17] Cowhage spicules contain a cysteine protease, mucuna in which is a pruritogenic substance that binds to proteinase-activated receptors, PAR 2 and PAR 4. [18] Cowhage-mediated pruritus may serve as an appropriate model to understand pruritus in atopic dermatitis [19] because PAR 2 receptors are elevated and involved in itch mediation in this condition. [20] Furthermore, cowhage-induced pruritus leads to concomitant pricking and burning sensations as seen in atopic dermatitis. [18]
It appears promising to pursue drugs capable of blocking the cowhage pathway for treatment of chronic pruritus and this pathway may serve as a new paradigm to test antipruritic agents in humans. [19]
It has been observed that non-histaminergic pruritogens may also mediate itch via toll-like receptor 7 (TLR-7) which is expressed by C-nerve fibers. [21]
Central Pathways of Itch
The C-nerve fibers synapse with lamina I neurons in the dorsal horn of spinal cord, then the secondary transmission neurons cross and enter into the opposite lateral STT to ascend to the thalamus. [12] Distinct lateral spinothalamic pathways conduct sensations from histamine and cowhage-sensitive neurons, respectively. A study by Sun et al. suggested that gastrin-releasing peptide receptor (GRPR) specifically mediates pruritus in the dorsal horn of the spinal cord but whether GRPR mediates other sensations could not be sufficiently excluded. [22] In a later study, GRPR+ neurons were selectively destroyed in mice by intrathecally injected bombesin-saporin. This led to a near complete loss in the scratching response to both histaminergic and non-histaminergic pruritogens, without affecting the pain behavior. [23] These results suggest that GRPR is specifically involved in the central mediation of itch at least in mice; however, this has not been confirmed in humans as yet.
SUPRASPINAL PROCESSING OF ITCH
Intracutaneously administered histamine-induced pruritus results in activation of multiple sites in the brain, which indicates that itch sensation is multidimensional and there is not a single itch center. [24] The areas of brain that are activated include anterior cingulate cortex, premotor area, primary somatosensory cortex parietal lobe (inferior), insular cortex, supplementary motor area, cerebellum and thalamus. [25],[26],[27],[28]
Activation of motor-associated areas correlates with the scratch response. A center in the distal medulla which in turn is under control of midbrain structures governs scratch movement. [29] The cerebellum might also have a function in coordinating the itch-scratch cycle. [30]
Mediators OF Itch
A bewildering myriad of mediators acting via a multitude of mechanisms are involved in pruritus.
Histamine
Histamine, despite being the most acknowledged mediator, plays a major part only in a few diseases like urticaria, insect bite reactions and mastocytosis. Dermal mast cell degranulation contributes to most of the histamine in the skin. Histamine acts on H1 receptors present on C-nerve fibers. H1 receptor is a G protein-coupled receptor attached to phospholipase Cί3 (PLCί). [31]
The role of histamine type 4 (H4) receptors expressed mainly on dendritic cells, mast cells and eosinophils is an exciting new field in our current understanding and management of pruritus. [32] These receptors are involved in allergic inflammation [33] and also in mediating pruritus. [34] This dual action makes H4 antagonists potential novel therapeutic agents for the management of allergic conditions like atopic dermatitis. [35] Prostaglandin (PG) E1 and E2 potentiate the pruritic action of histamine. [30]
Acetylcholine
Acetylcholine (Ach) is a neurotransmitter which binds to both muscarinic and nicotinic receptors. In mice, activation of the muscarinic M3 receptors causes pruritus. [36]
Studies revealed that intracutaneously injected acetylcholine induced pain [14] but it caused itch in atopic eczema. [37] Histamine-sensitive as well as histamine-insensitive C-nerve fibers are stimulated by acetylcholine. [7] Atopic dermatitis patients are more sensitive to acetycholine and less sensitive to histamine than normal subjects. [38] Since sympathetic nerves to eccrine sweat glands release acetylcholine, it is likely to be involved in the pruritus due to sweating seen in atopic dermatitis. [39]
Proteinases
The interest in the exact function of proteinases (like chymotrypsin, papain and trypsin) in itch has received renewed attention after studies revealed overexpression of chymotryptic enzyme within the stratum corneum in chronic itchy inflammatory skin diseases in mice. [40] Dermal mast cells produce tryptase and chymase which act through G protein-coupled receptor PAR 2 expressed on afferent C-neurons and keratinocytes. [18],[20],[41]
Elevated tryptase levels have been demonstrated in the lesional skin in comparison to non-lesional skin in patients of atopic dermatitis and the skin of normal controls. In addition, upregulation of PAR 2 receptors has been seen in atopic dermatitis. [20]
Activation of PAR 2 is also seen in itch due to kallikrein [42] and cathepsin as well as exogenous proteases including house-dust mite. [43] PAR 2 activation leads to the release of substance P as well as calcitonin gene-related peptide (CGRP) from nerve terminals which further leads to activation of CGRP receptors and neurokinin1 receptor, resulting in transmission of pruritic signals to the central nervous system. [41] However, intradermal injection of substance P causes itching but CGRP does not. [44]
It has been seen that both kallikreins and bradykinin can provoke pruritus by activating histamine-sensitive C-fibers. [14] Epidermal kallikrein-7 overexpression in mice leads to very intense itch behavior. [45] In addition, evidence suggests that pruritus induced by sodium deoxycholic acid in rodents is reduced by the use of bradykinin type-2 receptor antagonists. [46]
Cytokines
Interleukin 2
Interleukin 2 (IL2) is produced by activated T lymphocytes. Its role in pruritus is supported by the observation of redness and generalized pruritus following use of high dose recombinant IL2 for cancer treatment. [47] It is unclear as yet whether this process is mediated directly at the receptor level or indirectly through mast cells or endothelial cells. [5] Intradermal injection of IL2 has also been noted to induce pruritus. [48]
Interleukin 31
Interleukin 31 (IL31) is mainly produced by Th2 cells. IL31, through its regulation of both inflammation and pruritus, might be a link between the immunological and neurological systems. [5] Transgenic overexpression of IL31 has been found to evoke severe scratching and dermatitis in mice. [49] Decreased scratching behavior was observed on administering anti-IL31 antibodies in a mouse model of atopic dermatitis. [50] High levels of IL31 present in the skin of patients with atopic dermatitis and prurigo nodularis indicates its probable role in causing pruritus in these conditions. [51],[52]
IL31 binds a heterodimeric receptor composed of oncostatin M receptor and IL31 receptor A; but whether the pruritogenic effects are receptor mediated or indirect via keratinocytes is not completely known. [5] Targeting IL31 could serve as an innovative approach for treatment of itching in atopic eczema. [51]
Other Interleukins
Interleukin 6 levels are elevated in prurigo nodularis. [53] Increased expression of epidermal interleukin 4 (IL4) in transgenic mice results in an itchy inflammatory cutaneous disease similar to atopic dermatitis indicating the role of IL4 in pruritus and inflammation. [54] However, the role of TNF-alpha (TNF-α) and IL8 in pruritus remains unclear.
Interferon Gamma
It is quite clear that interferon gamma (IFN-γ) has beneficial antipruritic effects in atopic dermatitis but the mode of action is unknown. [5]
Substance P
Substance P is a neuro-peptide which acts mainly via neurokinin 1 (NK1) receptors. Intracutaneous injection of substance P produces itching as well as erythema and wheal. [55]
In atopic dermatitis, serum levels of substance P are elevated and correlate with severity of disease. [56] At physiological levels, substance P does not lead to histamine release from mast cells. [57] Application of substance P through a micro-dialysis probe leads to flare without wheals or itching. [58] However, it is clear that substance-P-expressing neurons are in direct communication with mast cells in the skin. [59]
At relevant concentrations, substance P plays a role in priming mast cells to other mast cell activators [60] resulting in release of TNF-α, [61] leukotriene B4, histamine and PG D2. Topical capsaicin which depletes substance P from cutaneous nerve terminals abolishes itch and pain. [62] In addition, a study by Costa et al.[63] indicates the role of NK1 receptor at the central level. NK1 receptor antagonist, apripetant, has been tried in Sezary syndrome for its antipruritic potential. [64]
Opioid Peptides
Pruritus caused by morphine is a well-recognized side effect in pain management. However, therapeutic opioid doses are inadequate to cause mast cell degranulation; therefore, opioid-induced pruritus is thought to be due to central mechanisms. But intradermal morphine does cause pruritus which can be inhibited by topical pre-treatment with the potent H1 antihistamine doxepine [65] but only partially inhibited by μ receptor antagonist naloxone suggesting both mast cell degranulation and activation of opioid receptors, both central and peripheral, are the possible mechanism of pruritus. This also suggests that histamine release by morphine is independent of μ receptor agonism. [66] Furthermore, intraspinal μ-opioid agonists induce segmental pruritus. [67] It has been observed that itch induced by intraspinal morphine can be inhibited by μreceptor antagonists [68] but is unaffected by antihistamines. [69] Thus, at the spinal level, opioid-induced itch is mediated by μ receptors. It has been seen that stimulating κ-opioid receptor blocks the effects of μ-receptor agonists. [70] Therefore, the generalized itch induced by opioids could possibly be an outcome of imbalance of μ receptor- and κ-receptor systems. [30] Opioid peptides have been used in the treatment of various disorders associated with pruritus. For example, μ-receptor antagonists naloxone and naltrexone have been effectively used in resistant itch associated with uremia and cholestasis; [71],[72] while κ receptor agonists such as nalfurafine have been effectively used in treating severe pruritus in patients undergoing hemodialysis. [73]
Transient Receptor Potential Channels in Pruritus
In general, transient receptor potential (TRP) molecules are calcium-permeable channels which sense temperature, osmotic and mechanical changes.
TRPV1 is present on nociceptive C-neurons and serves and is acted upon by capsaicin present in chilli pepper. In addition, it can be activated by various endogenous substances called endovanilloids. It has also been seen that TRP vanilloid 1 (TRPV1) is expressed on non-neuronal cells like keratinocytes. [74] Pruritogenic endovanilloids activate TRPV1 to initiate pruritus via a complex bidirectional intercellular system but repeated application of topical vanilloids desensitizes TRPV1-mediated signaling resulting in termination of itch. [75]
TRPs can be vital in the pathways of itch transmission. For example, it has been seen that sensory neuronal activation by histamine and PAR2 receptor activation also involves the activation/sensitization of TRPV1. [76] Accumulated evidence strongly suggests that TRPV1 is a fundamental integrating element in various pruritic and pain pathways. [5]
TRPV1 expression is amplified in keratinocytes of prurigo nodularis and becomes normal following capsaicin treatment. [74] In addition, stimulation of TRPV1 channels releases multiple pruritoceptive mediators like interleukins and neuropeptides. [77]
Other TRPVs (like TRPV2, TRPV3 and TRPV4) are activated at specific temperatures. [78] In view of the comparable neuronal expression of TRPV3 and TRPV1, and the postulate that TRPV3 may form heteropolymeric structures with TRPV1 monomers, [79] it is likely that TRPV3 might be a regulator and/or co-transducer of TRPV1-mediated pruritus and pain. It is pertinent to note that eicosanoids which possibly act as pruritogenic compounds activating TRPV1 may also activate TRPV4. [80] An interesting study by Stokes et al.[81] showed mast cell degranulation upon thermal and physical activation of TRPV2. In addition, mast cells also express TRPV1 and TRPV4.
TRP melastatin 8 (TRPM8) is expressed selectively by C-type neurons and acts as a cold thermosensor between 8°C and -28°C. Menthol and its analogs as well as icilin stimulate TRPM8. [82],[83] Menthol has been used as a topical antipruritic agent with moderate efficacy while topical icilin reduced excoriations in an animal model. [84] Similar effects to icilin were seen with carboxamides on TRPM8 in vitro, producing cold sensations lasting up to 1 hour and these may serve as models for endogenous TRPM8 agonists. [5]
TRP ankyrin 1 (TRPA1), nociceptor is a recently discovered mediator of non-histaminergic pruritus activated by MRGPRs in mice. [85]
Prostaglandins
Prostaglandin′s (PGs) potentiate itching caused by histamine as well as other mediators of pruritus. [86] Pre-treatment with PGE1 significantly lowers the threshold of pruritus induced by histamine and papain in humans. [87] However, orally administered aspirin, a cyclooxygenase inhibitor, had no effect in relieving itching [88] with the notable exception of polycythemia vera [89] but topically used aspirin alleviates pruritus in patients with chronic localized itch. [90] Non-steroidal anti-inflammatory drugs (NSAIDS) can also relieve pruritus in a subset of HIV patients probably by blocking cytokine-induced PGE2 production. [91] NSAIDS, e.g., per rectal diclofenac may reduce pruritus in morphine-induced spinal analgesia. [92] Studies in mice suggest that thromboxane-A2 [93] and leukotriene B4 are involved in pruritus. [94]
Neurotrophins and Nerve Growth Factor
Despite lack of direct evidence linking neurotrophins and pruritus many studies clearly demonstrate their role in pruritus. Nerve growth factor (NGF) is overexpressed in prurigo nodularis [95] and its therapeutic administration is pruritogenic. [96] In atopic dermatitis, NGF is released by keratinocytes, mast cells and fibroblasts [97] and plasma levels of NGF are also elevated and correlate with disease activity. [98] NGF induces release of the pruritogenic mediator tryptase. [97] In addition, expression of neurotrophin 4 is elevated in the cutaneous lesions of patients having atopic dermatitis and prurigo nodularis. [99]
Endocannabinoids and Cannabinoid Receptors
Studies have suggested that cannabinoid receptors are expressed on skin nerve fibers and may have a role in pruritus. For instance, cannabinoid receptor (CB1) agonist HU210 diminishes histamine-induced excitation of nerve fibers and thereby reduces itching. [100] This suggests CB1 signaling may be involved in initiation of itch. Furthermore, cannabinoids (like CB agonist anandamide) also activate the TRPV1 pathway. [101]
Serotonin (5-Hydroxytryptamine; 5HT)
Intradermal injection of serotonin in humans is mildly pruritogenic. [102] It causes histamine release from dermal mast cells. [103] Ondansetron a 5HT3-receptor antagonist alleviates itch secondary to opioid use, [104] suggesting a central mechanism as well. However, serotonin 3 receptor antagonists failed to relieve pruritus in renal failure. [105]
Calcitonin Gene-Related Peptide
Many neurons of the dorsal root ganglion co-express substance P (SP), CGRP, and PAR2. [41] CGRP plays a modulatory role in inflammation and pruritus. [106] It would seem that CGRP has an inhibitory effect on substance P-induced itching as it prolongs itch latency following injection, [107] but increased levels of CGRP are seen in atopic dermatitis, nummular eczema [108] and prurigo nodularis. [109] Like substance P, CGRP-mediated itch may result from mast cell activation. [110]
Corticotropin-Releasing Hormone
Corticotropin-releasing hormone (CRH) and its analog, urocortin, lead to histamine release upon intradermal injection. [111] Various mediators of pruritus are released in response to CRH probably via CRH-R. [112] CRH is also involved in mast cell degranulation occurring during periods of acute stress. [113]
The exact function that leukotrienes play in itch is unclear. Intradermal injection of leukotriene B4 provoked scratching in a mouse model. [114] In addition, studies have reported the use of leukotriene receptor antagonists zileuton and zafirlukast for their antipruritic action in atopic dermatitis. [115],[116]
Other Mediators
Intradermal injection of vasoactive intestinal polypeptide (VIP) induces itch which is dose dependent. [117] Somatostatin has also been reported to cause histamine release. [118] Nitric oxide augments pruritus induced by substance P in mice. [119] Bombesin induces scratching in rats following intraventricular injection. [120] Pruritus induced by bombesin was relieved by opioid analgesics pointing toward association with the opioid system. [121]
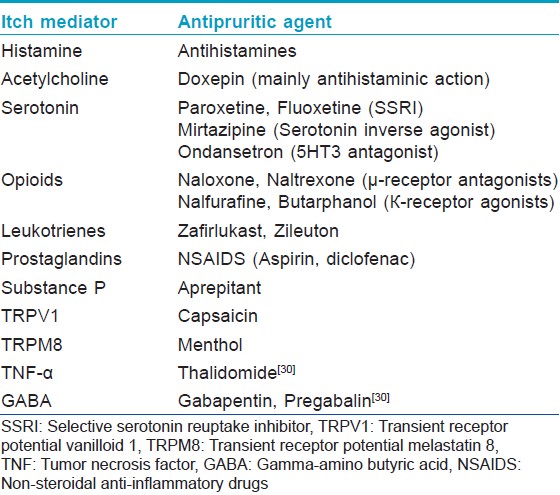
A summary of itch mediators and the corresponding possible antipruritic therapy is shown in the [Table - 1].
Conclusion
The complexity of the neurophysiological mechanisms causing pruritus as well as the extensive range of peripheral as well as central mediators of pruritus suggests that an effective antipruritic therapy would require a multipronged approach.
| 1. |
Hafenreffer S. Nosodochium, in quo cutis, eique adaerentium partium, affectusomnes, singulari methodo, et cognoscendi e curandi fidelisime traduntur. Ulmae(Westphalia) Kühnen; 1660. p. 98-102
[Google Scholar]
|
| 2. |
Rothman S. Physiology of itching. Physiol Rev1941;21:357-81.
[Google Scholar]
|
| 3. |
Savin J. How should we define itching? J Am Acad Dermatol1998;39:268-9.
[Google Scholar]
|
| 4. |
Yosipovitch G, Greaves MW. Definitions of itch. In: Yosipovitch G, Greaves MW, Fleischer AB, McGlone F, editors. Itch: Basic mechanisms and therapy. New York: Marcel Dekker; 2004. p. 1.
[Google Scholar]
|
| 5. |
Steinhoff M, Bienenstock J, Schmelz M, Maurer M, Wei E, BiroT. Neurophysiological, neuroimmunological, and neuroendocrine basis of pruritus. J Invest Dermatol 2006;126:1705-18.
[Google Scholar]
|
| 6. |
Kini SP, DeLong LK, Veledar E, McKenzie-Brown AM, Schaufele M, Chen SC. The impact of pruritus on quality of life: The skin equivalent of pain. Arch Dermatol 2011;147:1153-6.
[Google Scholar]
|
| 7. |
Twycross R, Greaves MW, Handwerker H, Jones EA, Libretto SE Szepietowski JC, et al. Itch: Scratching more than the surface. Q J Med 2003; 96:7-26.
[Google Scholar]
|
| 8. |
Ikoma A, Steinhoff M, Ständer S, Yosipovitch G, Schmelz M. Theneurobiology of itch. Nat Rev Neurosci 2006;7:535-47.
[Google Scholar]
|
| 9. |
Tuckett RP. Itch evoked by electrical stimulation of the skin. J Invest Dermatol 1982;79: 368-73.
[Google Scholar]
|
| 10. |
Schmelz M, Schmidt R, Bickel A, Handwerker HO, Torebjork HE. Specific C-receptors for itch in human skin. J Neurosci 1997;17:8003-8.
[Google Scholar]
|
| 11. |
Patel KN, Dong X. Itch: Cells, molecules, and circuits. ACS Chem Neurosci 2011;2:17-25.
[Google Scholar]
|
| 12. |
Andrew D, Craig AD.Spinothalamic lamina I neurons selectively sensitive to histamine: A central neural pathway for itch.Nat Neurosci 2001;4:72.
[Google Scholar]
|
| 13. |
Simone DA, Zhang X, Li J, Zhang JM, Honda CN, LaMotte RH, et al. Comparison of responses of primate spinothalamic neurons to pruritic and algogenic stimuli. J Neurophysiol 2004;91:213-22.
[Google Scholar]
|
| 14. |
Schmelz M, Schmidt R, Weidner C, Hilliges M, Torebjork HE, Handwerker HO.Chemical responsepatternofdifferentclasses of C-nociceptorstopruritogens and algogens. J Neurophysiol 2003;89:2441-8.
[Google Scholar]
|
| 15. |
Davidson S, Zhang X, Yoon CH, Khasabov SG, Simone DA, Giesler GJ Jr.The itch-producing agents histamine and cowhage activate separate populations of primate spinothalamic tract neurons. J Neurosci 2007;27:10007-14.
[Google Scholar]
|
| 16. |
Namer B, Carr R, Johanek LM, Schmelz M, Handwerker HO, Ringkamp M. Separate peripheral pathways for pruritus in man.J Neurophysiol 2008;100:2062-9.
[Google Scholar]
|
| 17. |
Johanek LM, Meyer RA, Hartke T, Hobelmann JG, Maine DN, LaMotte RH, et al.Psychophysical and physiological evidence for parallel afferent pathways mediating the sensation of itch. J Neurosci 2007;27:7490-7.
[Google Scholar]
|
| 18. |
Reddy VB, Luga AO, Shimada SG, LaMotte RH, Lerner EA. Cowhage- evoked itch is mediated by a novel cysteine protease: A ligand of protease-activated receptors. J Neurosci 2008;28:4331-5.
[Google Scholar]
|
| 19. |
Papoiu AD, Tey HL, Coghill RC, Wang H, Yosipovitch G. Cowhage-Induced Itch as an Experimental Model for Pruritus. A Comparative Study with Histamine-Induced Itch.PLoS One 2013;6:e17786.
[Google Scholar]
|
| 20. |
Steinhoff M, Neisius U, Ikoma A, Fartasch M, Heyer G, Skov PS, et al. Proteinase- activated receptor-2 mediates itch: A novel pathway for pruritus in human skin. J Neurosci 2003;23:6176-80.
[Google Scholar]
|
| 21. |
Liu T, Xu ZZ, Park CK, Berta T, Ji RR. Toll-like receptor 7 mediatespruritus. Nat Neurosci 2010;13:1460-2.
[Google Scholar]
|
| 22. |
Sun YG, Chen ZF. A gastrin-releasing peptide receptor mediates the itch sensation in the spinal cord. Nature 2007;448:700-3.
[Google Scholar]
|
| 23. |
Sun YG, Zhao ZQ, Meng XL, Jun Y, Yu XL, Feng CZ. Cellular basis of itchsensation. Science 2009;325:1531-4.
[Google Scholar]
|
| 24. |
Yosipovitch G, Greaves M, Schmelz M. Itch. Lancet 2003;36:690-4.
[Google Scholar]
|
| 25. |
Hsieh JC, Hägermark O, Ståhle-Bäckdahl M, Ericson K, Eriksson L, Stone-Elander S, et al. Urge to scratch represented in the human cerebral cortex during itch. J Neurophysiol1994;72:3004-38.
[Google Scholar]
|
| 26. |
Darsow U, Drzezega A, Frisch M, Munz F, Weilke F, Bartenstein P, et al. Processing of histamine induced itch in the human cerebral cortex: A correlation analysis withdermal reactions. J Invest Dermatol 2000;115:1029-33.
[Google Scholar]
|
| 27. |
Drzezga A, Darsow U, Treede RD, Siebner H, Frisch M, Munz F, et al. Central activation by histamine-induced itch: Analogies to pain processing: A correlationalanalysis of O-15 H2O positron emission tomography studies. Pain 2001;92:295-305.
[Google Scholar]
|
| 28. |
Mochizuki H, Tashiro M, Kano M, Sakurada Y, Itoh M, Yanai K. Imaging of central itch modulation in the human brain using positron emission tomography. Pain 2003;105:339-46.
[Google Scholar]
|
| 29. |
Ghez C. Voluntary movement. In: Kandel ER, Schwartz JH, editors. Principles of Neural Science. 2 nd ed. New York, NY: Elsevier; 1985:756-81
[Google Scholar]
|
| 30. |
Yosipovitch G, Papoiu A DP.Cutaneous Neurophysiology. In: Bolognia JL, Jorizzo JL, Schaffer JV, editors.Dermatology. 3 rd ed. New York: Elsevier; 2012:99-109.
[Google Scholar]
|
| 31. |
Han SK, Mancino V, Simon MI.Phospholipase Cβ 3 mediates the scratching response activated by the histamine H1 receptor on C-fiber nociceptive neurons. Neuron 2006;52:691-703.
[Google Scholar]
|
| 32. |
Huang JF, Thurmond R. The new biology of histamine receptors. Curr Allergy AsthmaRep 2008;8:21-7.
[Google Scholar]
|
| 33. |
Dunford PJ, O'Donnell N, Riley JP, Williams KN, Karlsson L, Thurmond RL, et al. The histamine H 4 receptor mediates allergic airway inflammation by regulating the activation of CD4+T cells. J Immunol 2006;176:7062-70.
et al. The histamine H 4 receptor mediates allergic airway inflammation by regulating the activation of CD4+T cells. J Immunol 2006;176:7062-70.'>[Google Scholar]
|
| 34. |
Bell JK, McQueen DS, Rees JL.Involvement of histamine H 4 and H 1 receptors in scratching induced by histamine receptor agonists in BalbC mice. Br J Pharmacol 2004;142:374-80.
[Google Scholar]
|
| 35. |
Cowden JM, Zhang M, Dunford PJ, Thurmond RL. The Histamine H 4 Receptor Mediates Inflammation and Pruritus in Th2-Dependent Dermal Inflammation.J Investig Dermatol 2010;130:1023-33.
[Google Scholar]
|
| 36. |
MiyamotoT, Nojima H, Kuraishi Y. Intradermalcholinergic agonists induce itch-associated responsevia M3 muscarinic acetylcholine receptors in mice. Jpn JPharmacol 2002;88:351-4.
[Google Scholar]
|
| 37. |
Heyer GR, Hornstein OP. Recent studies of cutaneous nociception in atopic and non-atopic subjects. J Dermatol 1999;26:77-86.
[Google Scholar]
|
| 38. |
Rukwied R, Lischetzki G, McGlone F, Heyer G, Schmelz M. Mast cell mediators other than histamine induce pruritus in atopic dermatitis: A dermal microdialysis study. Br J Dermatol 2000;142:1-8.
[Google Scholar]
|
| 39. |
Ikoma A.Updated Neurophysiology of Itch. Biol Pharm Bull 2013;36:1235-40.
[Google Scholar]
|
| 40. |
Hansson L, Backman A, Ny A, Edlund M, Ekholm E, Ekstrand Hammarström B, et al. Epidermal overexpression of stratum corneum chymotryptic enzyme in mice: A model for chronic itchy dermatitis. J Invest Dermatol 2002;118:444-9.
[Google Scholar]
|
| 41. |
Steinhoff M, Vergnolle N, Young SH, Tognetto M, Amadesi S, Ennes HS, et al. Agonists of proteinase-activated receptor 2 induce inflammation by a neurogenic mechanism. Nat Med 2000;6:151-8.
[Google Scholar]
|
| 42. |
Cormia FE, Dougherty JW. Proteolytic activity in development of pain anditching: Cutaneous reactions to bradykinin and kallikrein. J Invest Dermatol1960;35:21-6.
[Google Scholar]
|
| 43. |
Hägermark O. Itch mediators. Semin Dermatol 1995;14:271-6.
[Google Scholar]
|
| 44. |
Wallengren J, Hakanson R. Effects of substance P, neurokinin A and calcitonin gene-related peptide in humanskin and their involvement in sensory nerve-mediated responses. Eur J Pharmacol 1987;143:267-73.
[Google Scholar]
|
| 45. |
Ny A, Egelrud T. Epidermal hyperproliferation and decreased skin barrier function in mice overexpressing stratum corneum chymotryptic enzyme Acta Dermatol Venereol 2004;84:18-22.
[Google Scholar]
|
| 46. |
Hayashi I, Majima M. Reduction of sodium deoxycholicacid-induced scratching behaviour by bradykinin B2 receptor antagonists. Br J Pharmacol 1999;126:197-204.
[Google Scholar]
|
| 47. |
Gaspari AA, Lotze MT, Rosenberg SA, Stern JB, Katz SI. Dermatologic changes associated with interleukin 2 administration. JAMA 1987;258:1624-9.
[Google Scholar]
|
| 48. |
Darsow U, Scharein E, Bromm B, Ring J. Skin testing of the pruritogenic activity of histamine and cytokines (interleukin-2 and tumour necrosis factor- alpha) at the dermal-epidermal junction. Br J Dermatol 1997;137:415-7.
[Google Scholar]
|
| 49. |
Dillon SR, Sprecher C, Hammond A, Bilsborough J, Rosenfield-Franklin M, Presnell SR, et al. Interleukin 31, a cytokine producedby activated T cells, induces dermatitis in mice. Nat Immunol 2004;5:752-60.
[Google Scholar]
|
| 50. |
Grimstad O, Sawanobori Y, Vestergaard C, Bilsborough J, Olsen UB, Grønhøj-Larsen C, et al. Anti-interleukin-31-antibodies ameliorate scratching behaviour in NC/Nga mice: A model of atopic dermatitis. Exp Dermatol 2009;18:35-43.
[Google Scholar]
|
| 51. |
Sonkoly E, Muller A, Lauerma AI, Pivarcsi A, Soto H, Kemeny L, et al. IL-31: A new linkbetween T cells and pruritus in atopic skininflammation. J Allergy Clin Immunol 2006;117:411-7.
[Google Scholar]
|
| 52. |
Neis MM, Peters B, Dreuw A, Wenzel J, Bieber T, Mauch C, et al. Enhanced expression levels of IL-31 correlate with IL-4 and IL-13 in atopic and allergic contact dermatitis. J Allergy Clin Immunol 2006;118:930-7.
[Google Scholar]
|
| 53. |
Nordlind K, Chin LB, Ahmed AA, Brakenhoff J, Theodorssson E, Liden S.Immunohistochemical localization of interleukin-6-likeimmunoreactivity to peripheral nerve-like structures innormal and inflamed human skin. Arch Dermatol Res1996;288:431-5.
[Google Scholar]
|
| 54. |
Chan LS, Robinson N, Xu L. Expression of interleukin-4 in the epidermis oftransgenic mice results in a pruritic inflammatory skin disease: An experimental animal model to study atopic dermatitis. J Invest Dermatol 2001;117:977-83.
[Google Scholar]
|
| 55. |
Jorizzo JL, Coutts A, Greaves MW. Vascular responses of human skin to injection of substance P and mechanism of action. Eur J Pharmacol 1983;87:67-76.
[Google Scholar]
|
| 56. |
Toyoda M, Nakamura M, Makino T, Hino T, Kaqoura M, Morohashi M. Nerve growth factor and substance P are useful plasma markers ofdisease activity in atopic dermatitis. Br J Dermatol 2002;147:71-9.
[Google Scholar]
|
| 57. |
Schmelz M, Zeck S, Raithel M, Rukwied R. Mast celltryptase in dermal neurogenic inflammation. Clin ExpAllergy 1999;29:695-702.
[Google Scholar]
|
| 58. |
Weidner C, Klede M, Rukwied R, Lischetzki G, Neisius U, Skov PS, et al. Acute effects ofsubstance P and calcitonin gene-related peptide in humanskin: A microdialysis study. J Invest Dermatol 2000;115:1015-20.
[Google Scholar]
|
| 59. |
Suzuki R, Furuno T, McKay DM, Wolvers D, Tashima R, Nakanishi M, et al. Direct neurite-mast cell communicationin vitro occurs via the neuropeptide substance P. J Immunol1999;163:2410-5.
[Google Scholar]
|
| 60. |
Shanahan F, Denburg JA, Fox J, Bienenstock J, Befus D. Mast cell heterogeneity: Effects of neuroentericpeptides on histamine release. J Immunol1985;135:1331-7.
[Google Scholar]
|
| 61. |
Cocchiara R, Lampiasi N, Albeggiani G, Bongiovanni A, Azzolina A, Geraci D. Mast cellproduction of TNF-alpha induced by substance Pevidence for a modulatory role of substanceP-antagonists. J Neuroimmunol 1999;101:128-36.
[Google Scholar]
|
| 62. |
Nolano M, Simone DA, Wendelschafer-Crabb G, Johnson T, Hazen E, Kennedy WR. Topical capsaicin inhumans: Parallel loss of epidermal nerve fibres and painsensation. Pain 1999;81:135-45.
[Google Scholar]
|
| 63. |
Costa SK, Starr A, Hyslop S, Gilmore D, Bran SD. How importantare NK1 receptors for influencing microvascular inflammation and itch in the skin? Studies using Phoneutria nigriventorvenom.Vascul Pharmacol 2006;45:209-14.
[Google Scholar]
|
| 64. |
Duval A, Dubertret L. Aprepitant as an antipruritic agent? N Engl J Med 2009;361:1415-6.
[Google Scholar]
|
| 65. |
Heyer G, Dotzer M, Diepgen TL, Handwerker HO. Opiateand H1 antagonist effects on histamine inducedpruritus and alloknesis. Pain 1997;73:239-43.
[Google Scholar]
|
| 66. |
Blunk JA, Schmelz M, Zeck S, Skov P, Likar R, Koppert WOpioid-inducedmastcell activation and vascular responses is not mediated by μ-opioid receptors: An in vivomicrodialysisstudyinhumanskin. Anesth Analg 2004;98:364-70.
[Google Scholar]
|
| 67. |
Onigbogi O, Ajayi AA, Ukponmwan OE. Mechanisms of chloroquine-inducedbody scratchingbehaviorinrats: Evidenceof involvement of endogenous opioid peptides. Pharmacol Biochem Behav 2000;65:333-7.
[Google Scholar]
|
| 68. |
Saiah M, Borgeat A, Wilder-Smith OH, Rifat K, Suter PM.Epidural morphine induced pruritus: Propofol vs naloxone. Anesth Analg 1994;78:1110-3.
[Google Scholar]
|
| 69. |
Krause L, Shuster S.Mechanism of action of anti pruritic drugs.Br Med J (Clin Res Ed) 1983;287:1199-200.
[Google Scholar]
|
| 70. |
Umeuchi H, Togashi Y, Honda T, Nakao K, Okano K, Tanaka T, et al. Involvement ofcentral mu-opioid system in the scratching behavior inmice and the suppression of it by the activation ofkappa-opioid system. Eur J Pharmacol 2003;477:29-35.
[Google Scholar]
|
| 71. |
Peer G, Kivity S, Agami O, Fireman E, Silverberg D, Blum M, et al. Randomised crossover trial of naltrexone in uraemic pruritus.Lancet 1996;348:1552-4.
[Google Scholar]
|
| 72. |
Bergasa NV. The pruritus of cholestasis. J Hepatol 2005;43:1078-88.
[Google Scholar]
|
| 73. |
Kumagai H, Ebata T, Takamori K, Muramatsu T, Nakamoto H, SuzukiH. Effect of a novel kappa-receptor agonist, nalfurafine hydrochloride, on severe itch in 337 haemodialysis patients: A Phase III, randomized, double-blind, placebo-controlled study. Nephrol Dial Transplant2010;25:1251-7.
[Google Scholar]
|
| 74. |
Ständer S, Moormann C, Schumacher M, Buddenkotte J, Artuc M, Shpacovitch V, et al. Expression of vanilloid receptor subtype 1 in cutaneous sensory nerve fibers, mast cells, and epithelial cells of appendagestructures. Exp Dermatol 2004;13:129-39.
[Google Scholar]
|
| 75. |
Paus R, Schmelz M, Bíró T, Steinhoff M.Frontiers in pruritus research: Scratching the brain for more effective itch therapy . J Clin Invest2006;116:1174-85.
[Google Scholar]
|
| 76. |
Amadesi S, Nie J, Vergnolle N, Cottrell GS, Grady EF, TrevisaniM, et al. Protease- activated receptor 2 sensitizes the capsaicin receptortransientreceptorpotentialvanilloidreceptor1to inducehyperalgesia. J Neurosci 2004;24:4300-12.
[Google Scholar]
|
| 77. |
Southall MD, Li T, Gharibova LS, Pei Y, Nicol GD, Travers JB. Activation of epidermal vanilloid receptor-1 induces release of proinflammatory mediators in human keratinocytes.J Pharmacol Exp Ther 2003;304:217-22.
[Google Scholar]
|
| 78. |
Peier AM, Reeve AJ, Andersson DA, Moqrich A, Earley TJ, Hergarden AC, et al. Aheat-sensitive TRP channel expressed inkeratinocytes. Science 2002;296:2046-9.
[Google Scholar]
|
| 79. |
Smith GD, Gunthorpe MJ, Kelsell RE, Heyes PD, Reilly P, Facer P, et al.TRPV3 is a temperaturesensitivevanilloid receptor-like protein. Nature2002;418:186-90.
[Google Scholar]
|
| 80. |
Watanabe H, Vriens J, Prenen J, Droogmans G, Voets T, Nilius B. Anandamide and arachidonic acid use epoxyeicosatrienoic acids toactivate TRPV4 channels. Nature 2003;424:434-8.
[Google Scholar]
|
| 81. |
Stokes AJ, Shimoda LM, Koblan-Huberson M, Adra CN, Turner H. A TRPV2-PKAsignaling module for transduction of physicalstimuli in mast cells. J Exp Med 2004;200:137-47.
[Google Scholar]
|
| 82. |
McKemy DD, Neuhausser WM, Julius D.Identification of a cold receptor reveals ageneral role for TRP channels in thermosensation.Nature2002;416:52-8.
[Google Scholar]
|
| 83. |
Peier AM, Moqrich A, Hergarden AC, Reeve AJ, Andersson DA, Story GM, et al. ATRP channel that senses cold stimuli andmenthol. Cell 2002;108:705-15.
[Google Scholar]
|
| 84. |
Biro T, Ko MC, Bromm B, Wie ET, Bigliardi P, Siebenhaar F, et al. How best to fight that nastyitch from new insights into the neuroimmunological, neuroendocrine, and neurophysiologicalbases of pruritus to novel therapeutic approaches.ExpDermatol2005;14:225-40.
[Google Scholar]
|
| 85. |
Wilson SR, Gerhold KA, Bifolck-Fisher A, Liu Q, Patel KN, Dong X, et al. TRPA1 isrequired for histamine-independent, Mas-related Gprotein-coupled receptor-mediated itch. Nat Neurosci2011;14:595-602.
[Google Scholar]
|
| 86. |
Hagermark O, Strandberg K. Pruritogenic activity ofprostaglandin E2. Acta Derm Venereol 1977;57:37-43.
[Google Scholar]
|
| 87. |
Lovell CR, Burton PA, Duncan EH, Burton JL. Prostaglandins and pruritus. Br J Dermatol 1976;94:273-5.
[Google Scholar]
|
| 88. |
Daly BM, Shuster S. Effect of aspirin on pruritus. BMJ 1986;293:907.
[Google Scholar]
|
| 89. |
Jackson N, Burt D, Crocker J, Boughton B. Skin mastcells in polycythaemia vera: Relationship to the pathogenesis and treatment of pruritus. Br J Dermatol 1987;116:21-9.
[Google Scholar]
|
| 90. |
Yosipovitch G, Sugeng MW, Chan YH, Goon A, Ngim S, Goh CL. The effect of topically applied aspirinon localized circumscribed neurodermatitis. J Am Acad Dermatol2001;45:910-3.
[Google Scholar]
|
| 91. |
Smith KJ, Skelton HG, Yeager J. Pruritus in HIV-1 disease: Therapy with drugs which may modulate thepattern of immune dysregulation. Dermatology 1997;195:353-8.
[Google Scholar]
|
| 92. |
Colbert S, O'Hanlon DM, Galvin S, Chambers F, Moriarty DC. The effect of rectal diclofenac on pruritusin patients receiving intrathecal morphine. Anaesthesia 1999;54:948-52.
. The effect of rectal diclofenac on pruritusin patients receiving intrathecal morphine. Anaesthesia 1999;54:948-52.'>[Google Scholar]
|
| 93. |
Anhoh T, Nishikawa Y, Yamaguchi-Myamoto T, Nojima H, Narumiya S, Kuraiushi Y. Thromboxane A2 inducesitch-associated responses through TP receptors in the skin in mice. J Invest Dermatol 2007;127:2042-7.
[Google Scholar]
|
| 94. |
Andoh T, Yageta Y, Takeshima H, Kuraishi Y. Intradermalnociceptin elicits itch-associated responses throughleukotriene B(4) in mice. J Invest Dermatol 2004;123:196-201.
[Google Scholar]
|
| 95. |
Johansson O, Liang Y, Emtestam L. Increased nerve growth factor- and tyrosine kinase A-like immunoreactivities in prurigo nodularis skin: An exploration of the cause of neurohyperplasia. Arch Dermatol Res 2002;293:614-9.
[Google Scholar]
|
| 96. |
Aloe L. Rita Levi-Montalcini: The discoveryof nerve growth factor and modern neurobiology.Trends Cell Biol2004;14:395-9.
[Google Scholar]
|
| 97. |
Groneberg DA, Serowka F, Peckenschneider N, Artuc M, Grutzkau A, Fischer A, et al. Gene expression and regulation of nervegrowth factor in atopic dermatitis mast cellsand the human mast cell line-1. J Neuroimmunol2005;161:87-92.
[Google Scholar]
|
| 98. |
Toyoda M, Nakamura M, Makino T, Hino T, Kagoura M, Morohashi M. Nerve growth factor and substance P are useful plasma markers of disease activity inatopic dermatitis. Br J Dermatol 2002;147:71-9.
[Google Scholar]
|
| 99. |
Grewe M, Vogelsang K, Ruzicka T, Stege H, Krutmann J. Neurotrophin-4 production by human epidermal keratinocytes: Increased expression in atopic dermatitis. J Invest Dermatol 2000;114:1108-12.
[Google Scholar]
|
| 100. |
Dvorak M, Watkinson A, McGlone F, Rukwied R. Histamine induced responses are attenuated by cannabinoid receptor agonist in human skin. Inflamm Res 2003;52:238-45.
[Google Scholar]
|
| 101. |
Van der Stelt M, Trevisani M, Vellanni V, Petrocellis LD, Moriello AS, Campi B, et al.Anandamide acts as anintracellular messenger amplifying Ca(2+) influx via TRPV1 channels. EMBO J 2005;24:3517-8.
[Google Scholar]
|
| 102. |
Weisshaar E, Ziethen B, Gollnick H. Can a serotonin type 3 (5-HT3) receptor antagonist reduce experimentally-induced itch? Inflamm Res 1997;46:412-6.
[Google Scholar]
|
| 103. |
Weisshaar E, Ziethen B, Rohl FW, Gollnick H. Theantipruritic effect of a 5HT3 receptor antagonist (tropisetron) is dependent on mast cell depletion: An experimental study. Exp Dermatol 1999;8:254-60.
[Google Scholar]
|
| 104. |
Borgeat A, Stimemann HR. Ondansetron is effective totreat spinal or epidural morphine-induced pruritus. Anesthesiology 1999;90:432-6.
[Google Scholar]
|
| 105. |
Murphy M, Reaich D, Pai P, Finn P, Carmichael AJ. A randomized, placebo-controlleddouble-blind trial of ondansetron in renal itch. Br J Dermatol 2003;148:314-7.
[Google Scholar]
|
| 106. |
BrainSD, GrantAD. Vascularactionsof calcitonin gene-related peptide and adrenomedullin. Physiol Rev 2004;84:903-34.
[Google Scholar]
|
| 107. |
Ekblom A, Lundeberg T, Wahlgren CF. Influence of calcitonin gene-related peptide on histamine- and substance P-induced itch, flare and weal in humans. SkinPharmacol 1993;6:215-22.
[Google Scholar]
|
| 108. |
Jarvikallio A, Harvima IT, Naukkarinen A. Mast cells, nerves and neuropeptides in atopic dermatitis and nummular eczema. Arch Dermatol Res 2003;295:2-7.
[Google Scholar]
|
| 109. |
Liang Y, Jacobi HH, Reimert CM, Haak-Frendscho M, Marcusson JA, Johansson O. CGRP-immunoreactive nerves in prurigo nodularis: An exploration ofneurogenic inflammation. J Cutan Pathol 2000;27:359-66.
[Google Scholar]
|
| 110. |
Weidner C, Klede M, Rukwied R, Lischetzki G, Neisius U, Skov PS, et al. Acute effects of substance P and calcitonin gene-related peptide in human skin - a microdialysisstudy. J Invest Dermatol 2000;115:1015-20.
[Google Scholar]
|
| 111. |
Theoharides TC, Singh LK, Boucher W, Pang X, Letourneau R, Webster E, et al. Corticotropin-releasing hormone induces skin mast cell degranulation and increased vascular permeability, a possible explanation for its proinflammatory effects. Endocrinology 1998;139:403-13.
[Google Scholar]
|
| 112. |
Cao J, Papadopoulou N, Kempuraj D, Boucher WS, Sugimoto K, Cetrulo CL, et al. Humanmast cells express corticotropin releasing hormone (CRH) receptors and CRH leads to selective secretion of vascular endothelial growth factor.J Immunol 2005;174:7665-75.
[Google Scholar]
|
| 113. |
Singh LK, Pang X, Alexacos N, Letourneau R, Theoharides TC. Acute immobilization stress triggers skin mast cell degranulation via corticotropin releasing hormone, neurotensin, and substance P: A link to neurogenic skin disorders. Brain Behav Immun 1999;13:225-39.
[Google Scholar]
|
| 114. |
Andoh T, Yageta Y, Takeshima H, Kuraishi Y. Intradermal nociceptin elicits itch-associated responses throughleukotriene B(4) in mice. J Invest Dermatol 2004;123:196-201.
[Google Scholar]
|
| 115. |
Zabawski EJ Jr, Kahn MA, Gregg LJ. Treatment of atopic dermatitis with zafirlukast. Dermatol Online J 1999;5:10.
[Google Scholar]
|
| 116. |
Woodmansee DP, Simon RA. A pilot study examining the role of zileuton in atopic dermatitis. Ann Allergy Asthma Immunol. 1999;83:548-52.
[Google Scholar]
|
| 117. |
Rukwied R, Heyer G. Cutaneous reactions and sensations after intracutaneous injection of vasoactive intestinal polypeptide and acetylcholine in atopic eczema patients and healthy controls. Arch Dermatol Res 1998;290:198-204.
[Google Scholar]
|
| 118. |
Church MK, el-Lati S, Caulfield JP. Neuropeptide-induced secretion from human skin mast cells. Int Arch Allergy Appl Immunol 1991;94:310-8.
[Google Scholar]
|
| 119. |
Andoh T, Kuraishi Y. Nitric oxide enhances substanceP-induced itch-associated responses in mice. Br JPharmacol 2003;138:202-8.
[Google Scholar]
|
| 120. |
Gmerek DE, Cowan A. Bombesin: A central mediator of pruritus? Br J Dermatol 1983;109:239.
[Google Scholar]
|
| 121. |
Gmerek DE, Cowan A. An animal model for preclinical screening of systemic antipruritic agents.J Pharmacol Methods 1983;10:107-12.
[Google Scholar]
|
Fulltext Views
14,460
PDF downloads
4,338





![[Table - 1]](#tbl_ijdvl_2014_80_2_106_129377_t1.jpg){kind=link}